永野真吾 研究内容
アナモックス菌が持つユニークな梯子状疎水基を持つラダラン脂質の生合成マシナリーの解明
 空気の80パーセントは分子状窒素であり、私たちの体にもタンパク質や核酸など窒素を含む物質が多く存在しています。これらの窒素化合物は主に微生物によって変換され地球上の窒素バランスが保たれています。硝化・脱窒・窒素固定など古くから知られている窒素循環に加えて、
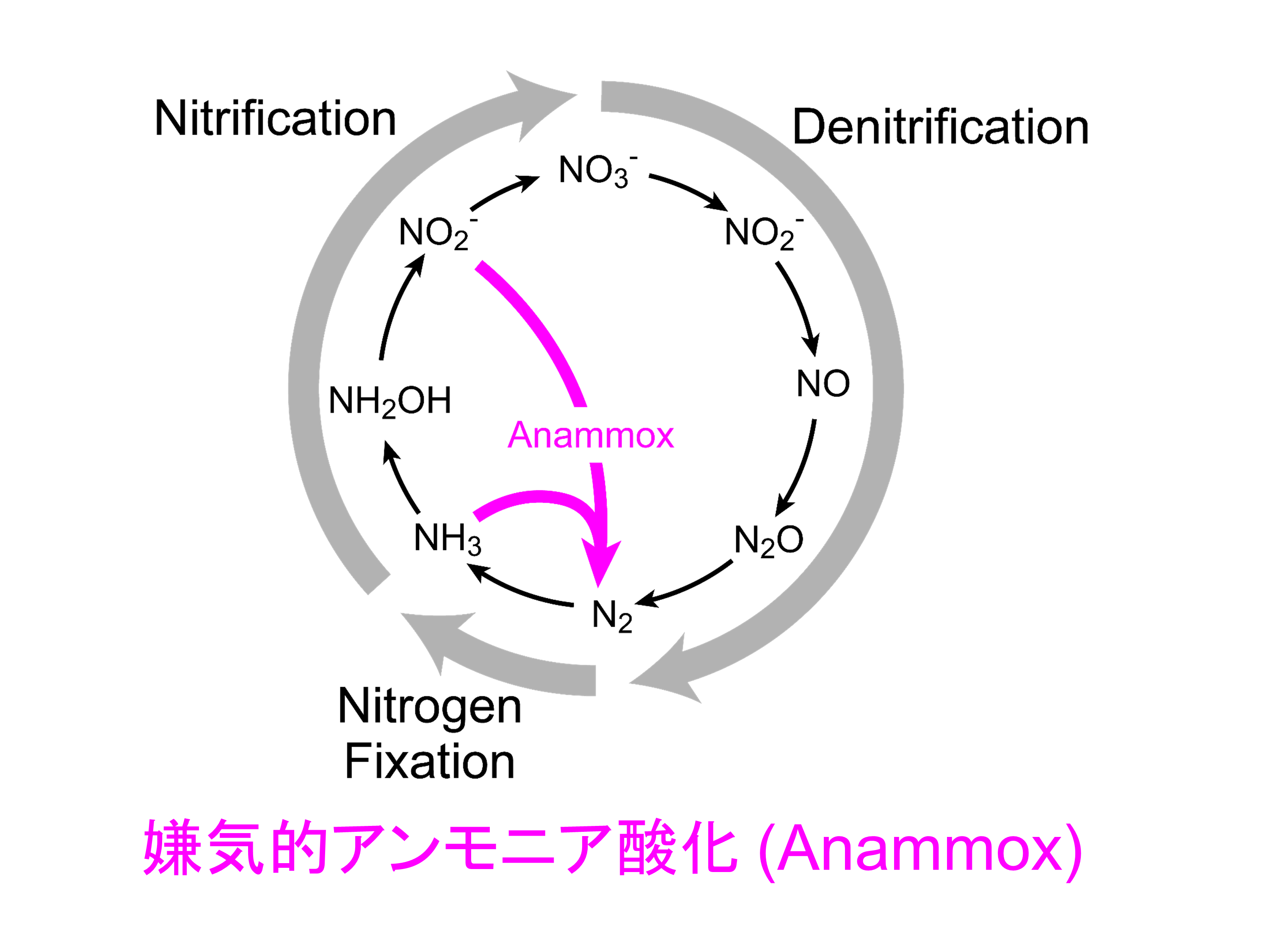
アンモニアと亜硝酸から分子状窒素を作り出すアナモックス反応が1990年代に発見されました。
空気の80パーセントは分子状窒素であり、私たちの体にもタンパク質や核酸など窒素を含む物質が多く存在しています。これらの窒素化合物は主に微生物によって変換され地球上の窒素バランスが保たれています。硝化・脱窒・窒素固定など古くから知られている窒素循環に加えて、
アンモニアと亜硝酸から分子状窒素を作り出すアナモックス反応が1990年代に発見されました。
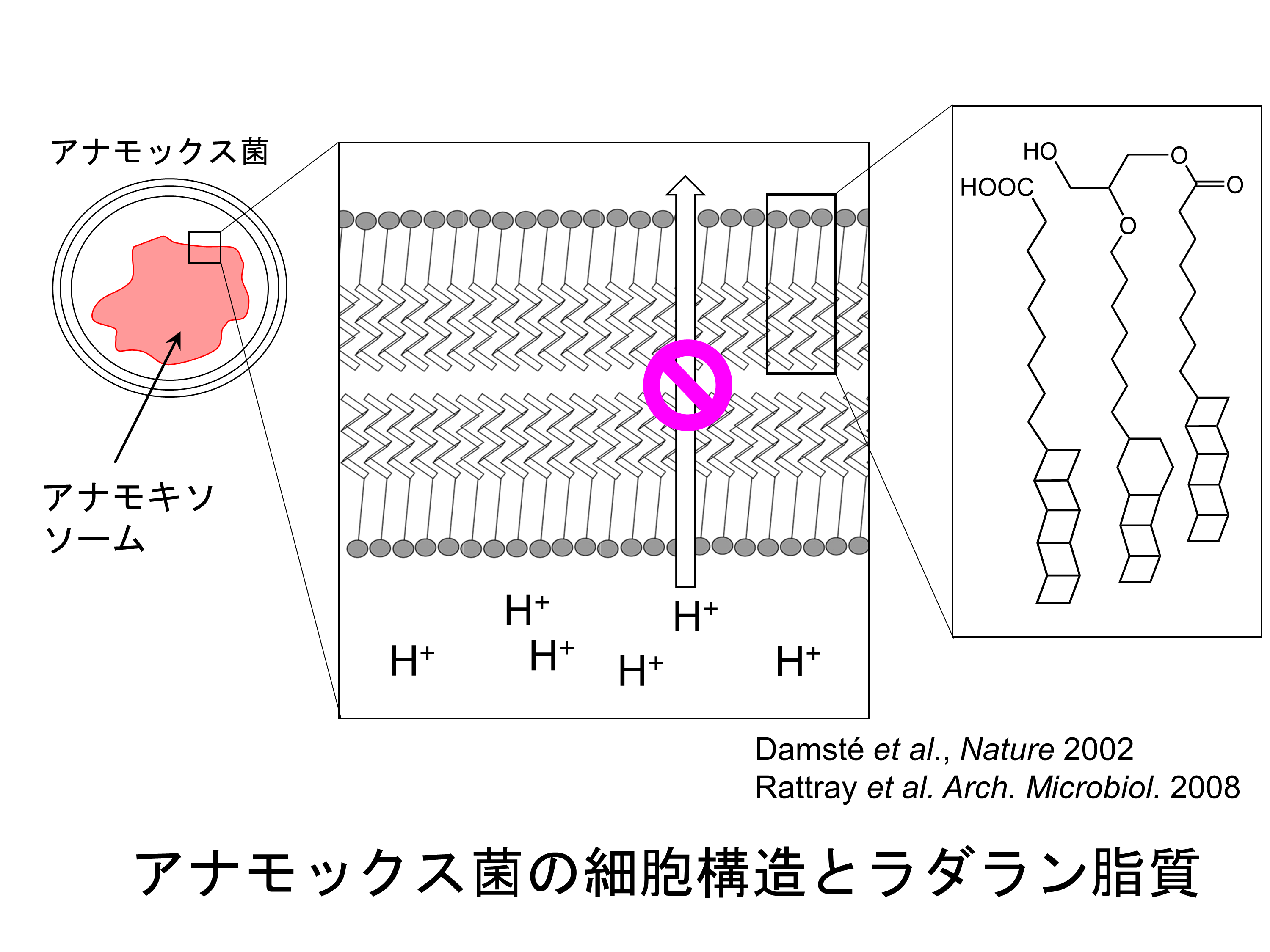
 この反応を行うアナモックス菌はきわめて増殖が遅いため、ATPの合成を駆動するプロトンの濃度勾配もゆっくりと形成されますが、通常の脂質二重膜ではわずかですがプロトンが漏れるため、ATPの合成効率が非常に悪くなってしまいます。アナモックス菌は、
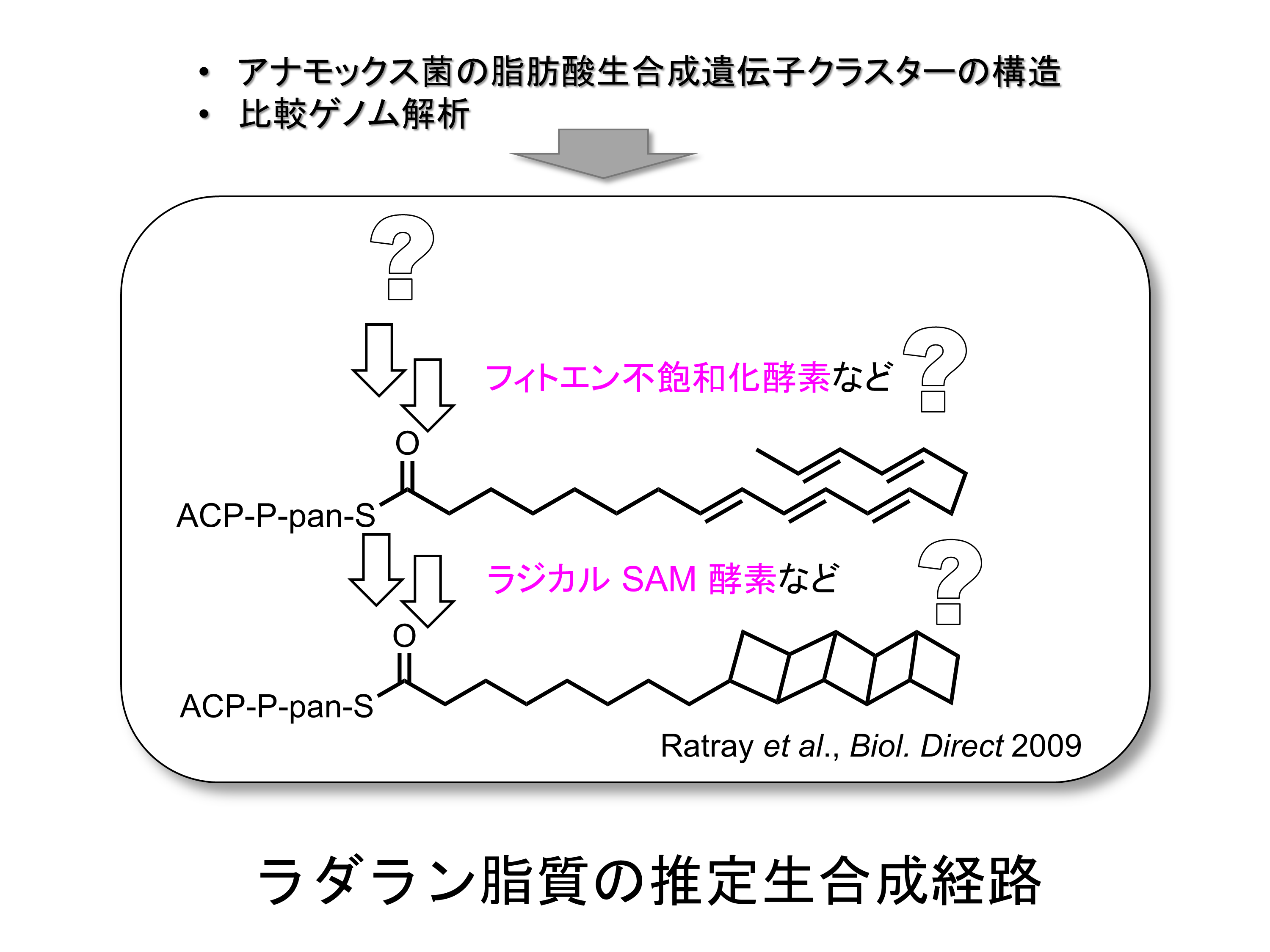
四員環が連なった梯子状疎水基を持つラダラン脂質を多く含む稠密な膜でプロトンの濃度勾配を維持していると考えられています。ラダラン脂質の生理的な役割に加えて、その極めてユニークな構造は多くの天然物生合成や有機化学の研究者の注目を集めました。また、梯子状疎水基には歪みとして大きなエネルギーが蓄積されているため、ラダラン脂質は高いエネルギー密度を持つバイオジェット燃料として応用が期待されています。ラダラン脂質の梯子構造を試験管内で合成することは非常に困難であるため、アナモックス菌がどうやってこの脂質を生合成しているのか、その仕組みに興味が持たれますが、いまだに大部分が謎のままです。Rattrayらはアナモックス菌のゲノムを詳細に解析し、長鎖飽和脂肪酸から多価不飽和脂肪酸が生合成され、これを前駆体としてラダラン脂質が産生される生合成経路を提案しました。私たちはこの仮説に基づき、ラダラン脂質の梯子構造の生合成中間体を探し出す研究や、梯子構造を構築する鍵反応を行うと推定されるいくつかの酵素の結晶構造解析などの研究を進めています。ラダラン脂質の梯子構造が作られる仕組みを明らかにすることができれば、複雑な炭素骨格を持ち有用な生物活性を持つ化合物を自在に設計できる基盤が作られると期待されます。さらに、地球温暖化ガスの増加を抑え環境負荷が小さいバイオ燃料の生産に結び付く可能性もあります。
この反応を行うアナモックス菌はきわめて増殖が遅いため、ATPの合成を駆動するプロトンの濃度勾配もゆっくりと形成されますが、通常の脂質二重膜ではわずかですがプロトンが漏れるため、ATPの合成効率が非常に悪くなってしまいます。アナモックス菌は、
四員環が連なった梯子状疎水基を持つラダラン脂質を多く含む稠密な膜でプロトンの濃度勾配を維持していると考えられています。ラダラン脂質の生理的な役割に加えて、その極めてユニークな構造は多くの天然物生合成や有機化学の研究者の注目を集めました。また、梯子状疎水基には歪みとして大きなエネルギーが蓄積されているため、ラダラン脂質は高いエネルギー密度を持つバイオジェット燃料として応用が期待されています。ラダラン脂質の梯子構造を試験管内で合成することは非常に困難であるため、アナモックス菌がどうやってこの脂質を生合成しているのか、その仕組みに興味が持たれますが、いまだに大部分が謎のままです。Rattrayらはアナモックス菌のゲノムを詳細に解析し、長鎖飽和脂肪酸から多価不飽和脂肪酸が生合成され、これを前駆体としてラダラン脂質が産生される生合成経路を提案しました。私たちはこの仮説に基づき、ラダラン脂質の梯子構造の生合成中間体を探し出す研究や、梯子構造を構築する鍵反応を行うと推定されるいくつかの酵素の結晶構造解析などの研究を進めています。ラダラン脂質の梯子構造が作られる仕組みを明らかにすることができれば、複雑な炭素骨格を持ち有用な生物活性を持つ化合物を自在に設計できる基盤が作られると期待されます。さらに、地球温暖化ガスの増加を抑え環境負荷が小さいバイオ燃料の生産に結び付く可能性もあります。
植物の成長ホルモン「ブラシノステロイド」の生合成に関わるシトクロムP450
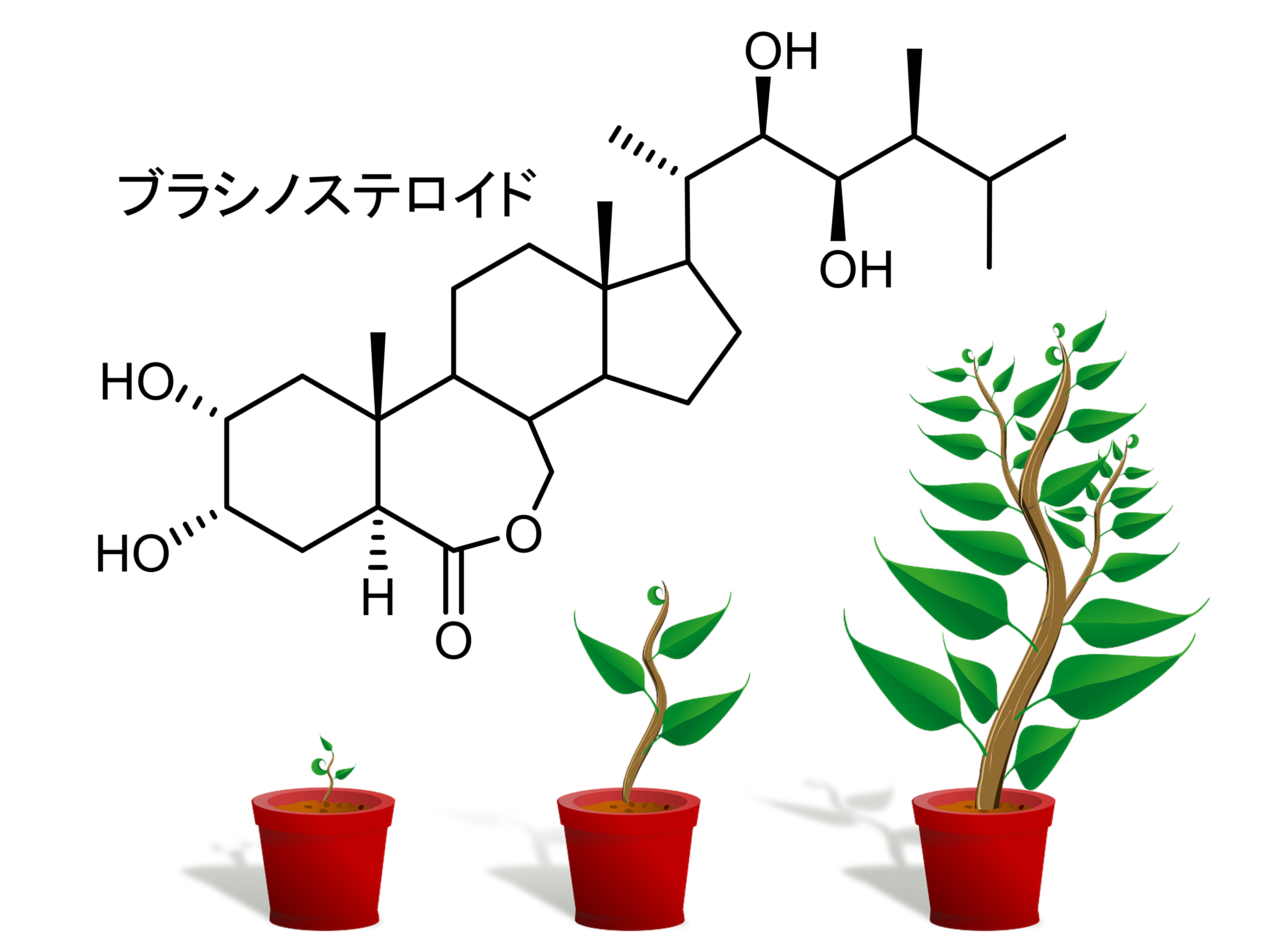 思春期のヒトの体内では成長ホルモンが多く分泌され、体が大きく成長します。また、テストステロンやエストラジオールなどのステロイドホルモンは性ホルモンとして機能し、男性と女性の体つきの違いを生じさせます。植物も
ブラシノステロイドと呼ばれるステロイドホルモンを作り出しており、植物の成長ホルモンとして機能しています。ブラシノステロイドはカンペステロールを前駆体として作り出されていますが、これらの過程で様々なシトクロムP450(P450)とよばれる酵素によって多くの酸素原子が添加されます。CYP90B1はシトクロムP450の一種であり、ブラシノステロイド生合成において最初の酸素添加反応を行い、このプロセスがブラシノステロイド生合成の速度を決める律速段階となっています。従って、CYP90B1はブラシノステロイドの生合成系の中で最も重要な酵素です。私たちの研究室では、CYP90B1の結晶構造を世界に先駆けて解析し、この酵素が酸素原子を添加するメカニズムを明らかにすることができました。この成果は国内外で大きな注目を集め、この構造を基に植物の成長を制御できる農薬として利用できる化合物の開発が期待されます。私たちはCYP90B1以外にもブラシノステロイド生合成で働いている重要なシトクロムP450の研究にも取り組んでいます。
思春期のヒトの体内では成長ホルモンが多く分泌され、体が大きく成長します。また、テストステロンやエストラジオールなどのステロイドホルモンは性ホルモンとして機能し、男性と女性の体つきの違いを生じさせます。植物も
ブラシノステロイドと呼ばれるステロイドホルモンを作り出しており、植物の成長ホルモンとして機能しています。ブラシノステロイドはカンペステロールを前駆体として作り出されていますが、これらの過程で様々なシトクロムP450(P450)とよばれる酵素によって多くの酸素原子が添加されます。CYP90B1はシトクロムP450の一種であり、ブラシノステロイド生合成において最初の酸素添加反応を行い、このプロセスがブラシノステロイド生合成の速度を決める律速段階となっています。従って、CYP90B1はブラシノステロイドの生合成系の中で最も重要な酵素です。私たちの研究室では、CYP90B1の結晶構造を世界に先駆けて解析し、この酵素が酸素原子を添加するメカニズムを明らかにすることができました。この成果は国内外で大きな注目を集め、この構造を基に植物の成長を制御できる農薬として利用できる化合物の開発が期待されます。私たちはCYP90B1以外にもブラシノステロイド生合成で働いている重要なシトクロムP450の研究にも取り組んでいます。
トマチン生合成の鍵酵素2-オキソグルタル酸依存性ジオキシゲナーゼ
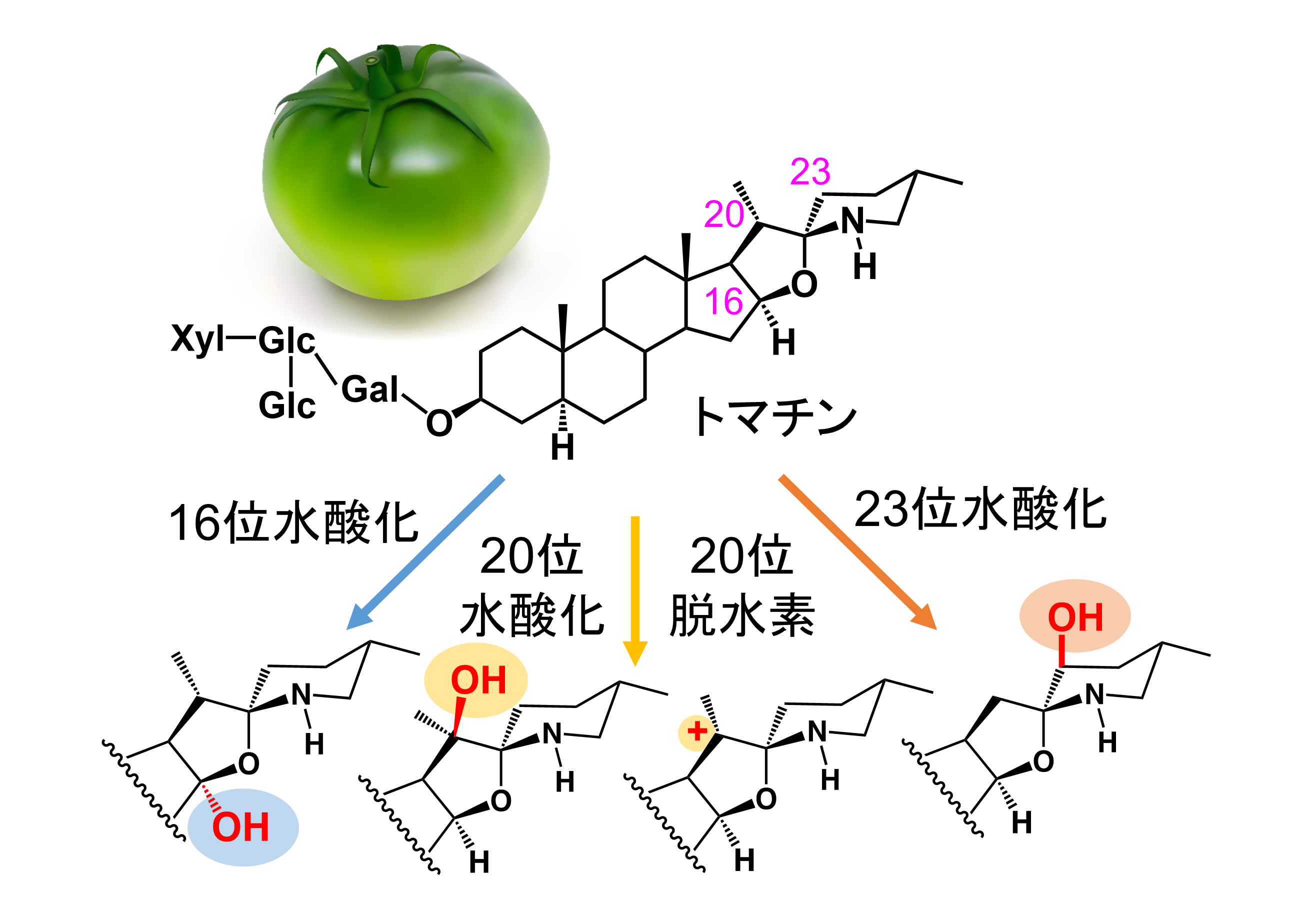 トマチンは未成熟のトマト果実に含まれる苦味物質であり、昆虫の忌避成分や土壌伝染病に対する抗菌物質として機能しています。この化合物は果実の成熟に伴って分解されますが、その初発反応も、クマリン生合成の鍵段階と同様に非ヘム鉄と2-オキソグルタル酸依存性のジオキシゲナーゼ (2OGD) が触媒しています。植物は、トマチンの様々な部位に対して水酸化や脱水素反応などを行い、多様なトマチン代謝のきわめて重要な役割が明らかになりつつあります。当研究室では、トマチンを共通の基質としながら、異なる部位の酸化反応を特異的に触媒するトマチン代謝2OGDのX線結晶構造解析を進めています。これらの構造から多様なトマチン代謝のメカニズムを解明します。
トマチンは未成熟のトマト果実に含まれる苦味物質であり、昆虫の忌避成分や土壌伝染病に対する抗菌物質として機能しています。この化合物は果実の成熟に伴って分解されますが、その初発反応も、クマリン生合成の鍵段階と同様に非ヘム鉄と2-オキソグルタル酸依存性のジオキシゲナーゼ (2OGD) が触媒しています。植物は、トマチンの様々な部位に対して水酸化や脱水素反応などを行い、多様なトマチン代謝のきわめて重要な役割が明らかになりつつあります。当研究室では、トマチンを共通の基質としながら、異なる部位の酸化反応を特異的に触媒するトマチン代謝2OGDのX線結晶構造解析を進めています。これらの構造から多様なトマチン代謝のメカニズムを解明します。
クマリン生合成の鍵酵素2-オキソグルタル酸依存性ジオキシゲナーゼ
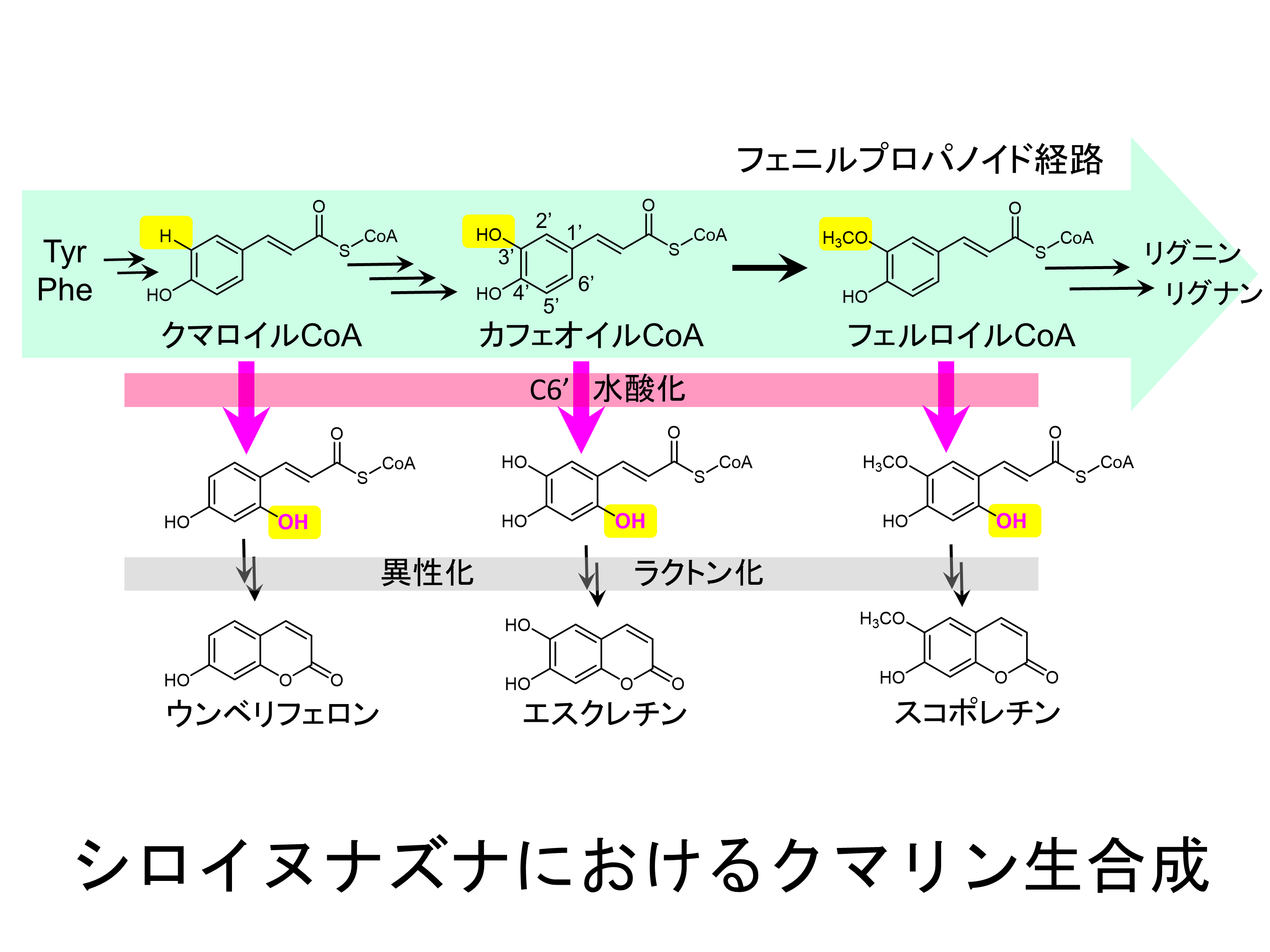 桜餅は独特の芳香を持ちますが、これは
クマリンと呼ばれる芳香族化合物が持つ香りです。植物はクマリンや類似の化合物を生産し、これらは植物病原菌に対する抗菌物質として働くことが知られています。クマリンは植物の中で桂皮酸類CoA体を出発物質として、C6’水酸化反応とその後の非酵素的な異性化と環化反応によって生合成されています。この反応の鍵段階である桂皮酸類CoA体の水酸化反応を触媒するのが非ヘム鉄と2-オキソグルタル酸を補因子に持つジオキシゲナーゼ (2OGD) です。クマリンの生合成に関与する2OGD (Cm 2OGD) は、様々な植物に存在することが知られており、それらは基質のわずかな構造の違いを認識し、異なる基質選択性を持っています。当研究室ではシロイヌナズナが持つクマリンの生合成に関わる2OGDのX線結晶構造解析、ドッキングシミュレーションや生化学的な解析から、多様なクマリン生合成の仕組みを明らかにする研究に取り組んでいます。
桜餅は独特の芳香を持ちますが、これは
クマリンと呼ばれる芳香族化合物が持つ香りです。植物はクマリンや類似の化合物を生産し、これらは植物病原菌に対する抗菌物質として働くことが知られています。クマリンは植物の中で桂皮酸類CoA体を出発物質として、C6’水酸化反応とその後の非酵素的な異性化と環化反応によって生合成されています。この反応の鍵段階である桂皮酸類CoA体の水酸化反応を触媒するのが非ヘム鉄と2-オキソグルタル酸を補因子に持つジオキシゲナーゼ (2OGD) です。クマリンの生合成に関与する2OGD (Cm 2OGD) は、様々な植物に存在することが知られており、それらは基質のわずかな構造の違いを認識し、異なる基質選択性を持っています。当研究室ではシロイヌナズナが持つクマリンの生合成に関わる2OGDのX線結晶構造解析、ドッキングシミュレーションや生化学的な解析から、多様なクマリン生合成の仕組みを明らかにする研究に取り組んでいます。
立体選択的なディールス・アルダー反応によって抗HIV活性を持つエキセチンやフォマセチンの分子骨格を構築する酵素
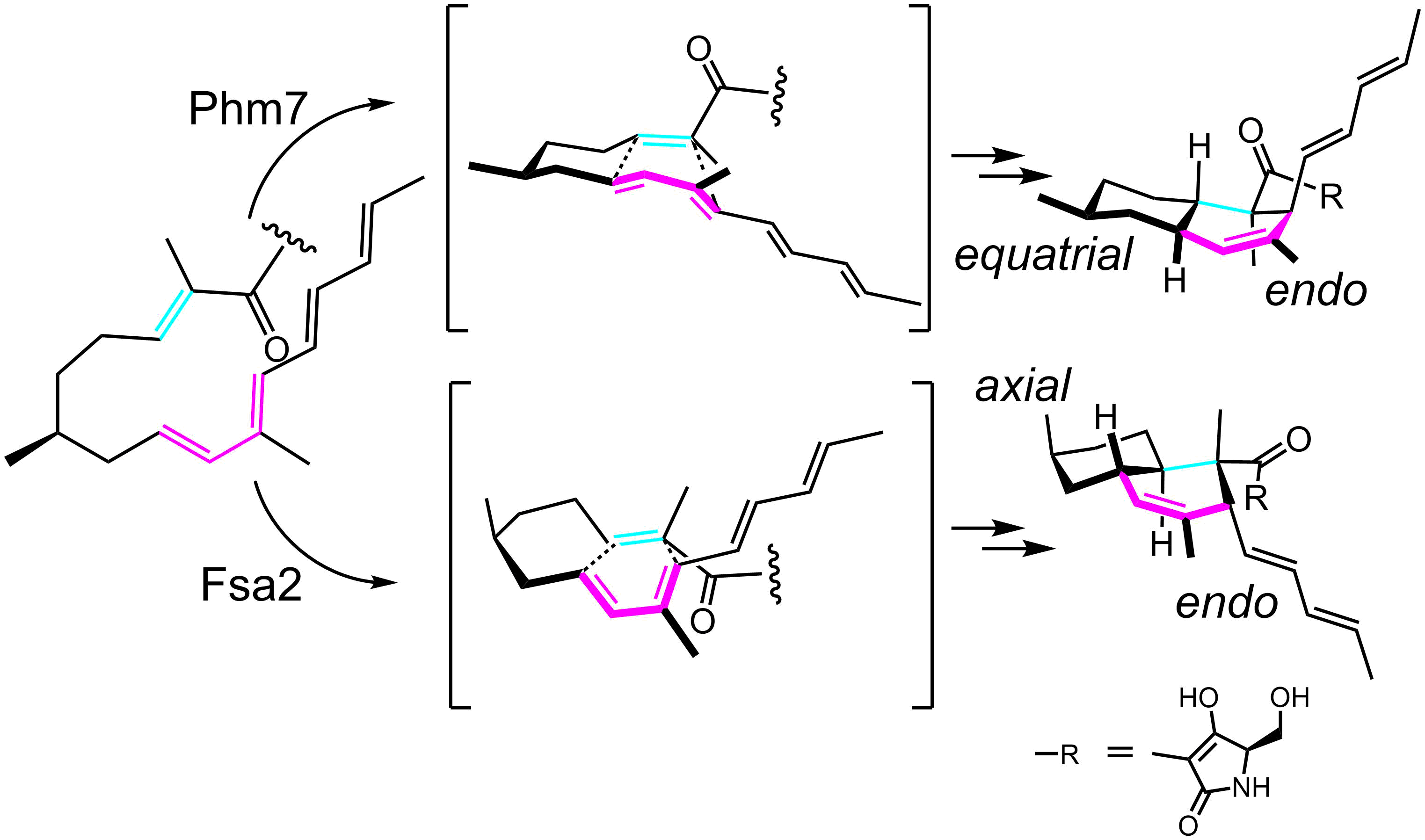 ディールス・アルダー反応は有機合成化学で有用な6員環形成反応であり、共役ジエンとアルケンから環化付加反応でシクロヘキセンが形成される。自然界にはディールス・アルダー反応を触媒する酵素が存在することが知られている。カビの一種Fusarium sp. FN080326は、抗がん活性を示すフサリセチンを作りだしますが、その生合成の中間体はフォマセチンと呼ばれる6員環を二つもつ化合物であり、この化合物は抗HIV活性を示すことから注目を集めています。フォマセチンの2つの6員環からなる骨格はデカリン骨格と呼ばれ、この骨格構造は図のように酵素Fsa2が触媒する分子内ディールス・アルダー反応によって構築されます。さらに、興味深いことにFsa2とアミノ酸配列が類似した酵素Phm7も(ほぼ)同じ基質に対して分子内ディールス・アルダー反応を触媒しますが、立体選択性が異なっています。私たちはこれらの酵素が立体選択的に分子内ディールス・アルダー反応を触媒するメカニズムの解明を、これらの酵素のX線結晶構造解析などにより進めています。
ディールス・アルダー反応は有機合成化学で有用な6員環形成反応であり、共役ジエンとアルケンから環化付加反応でシクロヘキセンが形成される。自然界にはディールス・アルダー反応を触媒する酵素が存在することが知られている。カビの一種Fusarium sp. FN080326は、抗がん活性を示すフサリセチンを作りだしますが、その生合成の中間体はフォマセチンと呼ばれる6員環を二つもつ化合物であり、この化合物は抗HIV活性を示すことから注目を集めています。フォマセチンの2つの6員環からなる骨格はデカリン骨格と呼ばれ、この骨格構造は図のように酵素Fsa2が触媒する分子内ディールス・アルダー反応によって構築されます。さらに、興味深いことにFsa2とアミノ酸配列が類似した酵素Phm7も(ほぼ)同じ基質に対して分子内ディールス・アルダー反応を触媒しますが、立体選択性が異なっています。私たちはこれらの酵素が立体選択的に分子内ディールス・アルダー反応を触媒するメカニズムの解明を、これらの酵素のX線結晶構造解析などにより進めています。
インドール環の位置選択的なプレニル化を行い、インドールアルカロイドの構造多様性を生み出す酵素
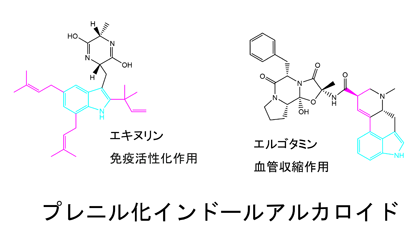
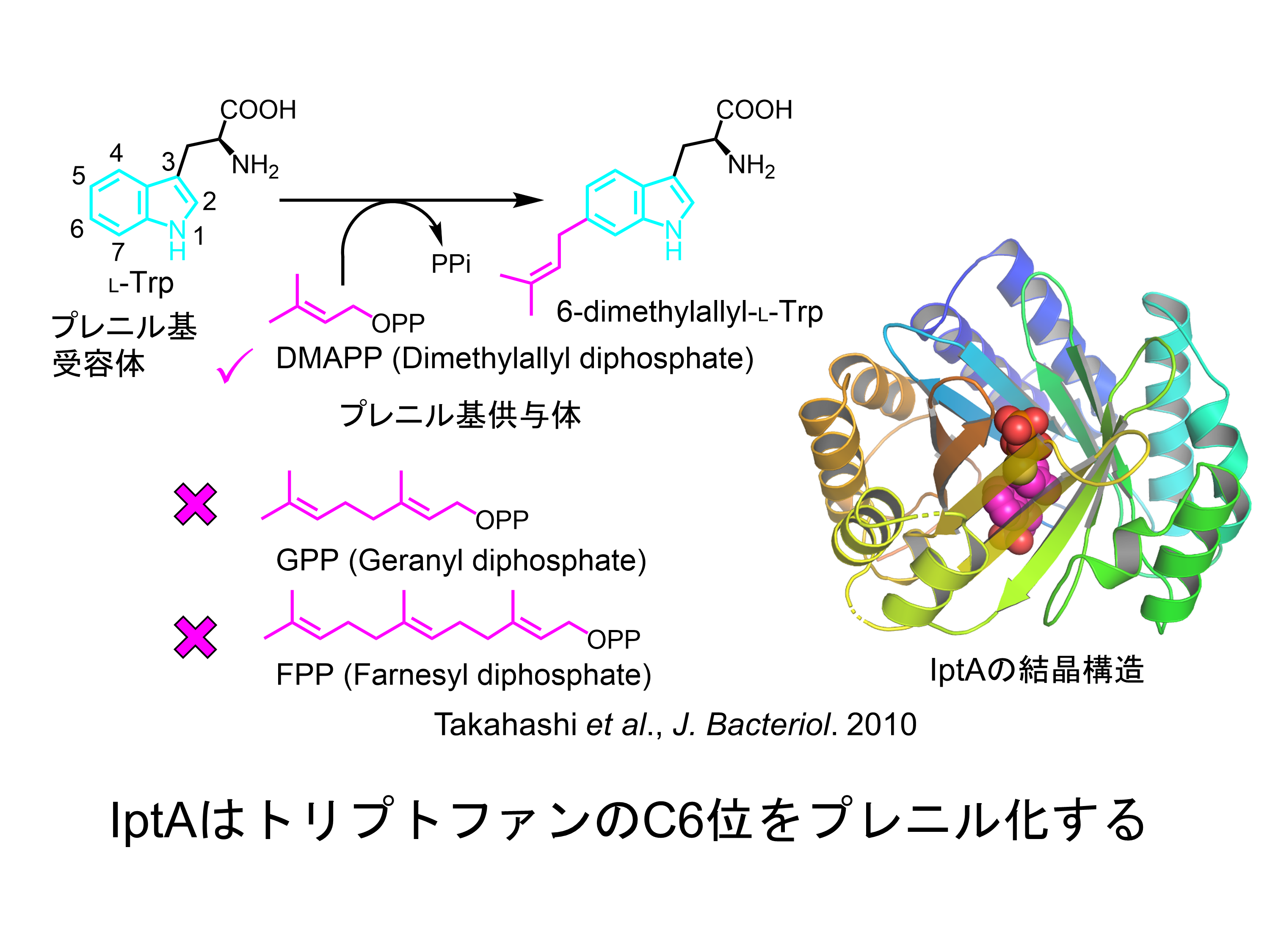
プレニル化されたインドール環を持つアルカロイドは主に植物や微生物によって作られ、多様な構造と生理作用をもつものが数多く知られています。例えばエキヌリンは免疫活性化作用を持ち、エルゴタミンは血管収縮作用を示します。これらの化合物でプレニル基はインドール環の側鎖、または分子骨格の一部として存在しています。これらのプレニル基はインドールプレニル化酵素によってトリプトファンやインドールを含む化合物に付加されます。プレニル化によって疎水性が高められ、新たな骨格構築の足場となります。これまでに多くのインドールプレニル化酵素が生化学的に解析され、インドール環のすべての位置に対してプレニル化を行う酵素が存在することが知られています。私たちの共同研究者、理化学研究所の高橋博士は、放線菌Streptomyces sp. SN-593からトリプトファンのインドール環の6位を位置選択的にプレニル化する酵素IptAを初めて発見しました。この酵素は様々な置換基が付加されたトリプトファン誘導体に対してもプレニル化を行うことができる、すなわちプレニル基受容体にたいして寛容性を示すことが特徴の一つです。一方、この酵素はゲラニル二リン酸やファルネシル二リン酸など鎖長が長いプレニル基を用いてプレニル化することはできません。つまり、プレニル基供与体に対しては非寛容な酵素です。私たちは、IptAの位置選択的なプレニル化、プレニル基受容体に対する寛容性、そしてプレニル基供与体に対する不寛容性が現れる仕組みを明らかにするために、プレニル基受容体、プレニル基供与体アナログがIptAに結合した三者複合体のX線結晶構造解析を行っています。また、近年様々な生物活性を持つペプチドが中分子医薬品として注目されていますが、IptAはトリプトファンを含むペプチドもプレニル化することができるため、ペプチドの生理機能の拡張に有用な酵素として活用が期待されます。